表观遗传调控机制是牙周病发病和发展的重要危险因素。关于牙周病的炎症方面,组蛋白尾部氨基酸的翻译后修饰(PTMs)已被研究。组蛋白的PTMs包括乙酰化和甲基化等,它们以环境依赖性的方式调节转录活性。虽然组蛋白乙酰转移酶(HATs)对乙酰基的附着通常与染色质结构的开放和基因表达的有利环境有关,但组蛋白甲基化的影响取决于被修饰的氨基酸和这些甲基的丰度。例如,组蛋白H3(H3)第27位赖氨酸(K27)的三甲基化修饰是一类重要的转录抑制性翻译后修饰,在生物进程的各个方面都发挥着重要作用。
血清长链脂肪酸水平升高,如饱和脂肪酸棕榈酸(PA)通常与肥胖有关,此外,在高脂血症条件下,PA具有促炎特征。一些研究报告了这两种疾病之间的相互关系,例如,在喂食高脂饮食的小鼠中观察到对革兰阴性厌氧菌牙龈卟啉单胞菌(P. gingivalis)感染的延迟反应。牙龈卟啉单胞菌已被描述为影响口腔健康和疾病的关键病原体,可能是由于其逃避宿主免疫反应的独特能力。
尽管这两种疾病中的炎症过程都增加,但它们在受到机械力(例如创伤、咀嚼或正畸牙齿运动期间施加的力)时对牙周膜(PdL)炎症反应的伴随影响仍然研究不足。PdL是牙齿和牙槽骨之间的结缔组织,其最丰富的细胞是PdL成纤维细胞(PdLFs),以时间和空间方式调节对压缩力的短暂无菌炎症反应。
研究最近报道,PA刺激的人PdL成纤维细胞(HPdLFs)对牙龈卟啉单胞菌脂多糖(LPS)和压缩力(2 g/cm²)的同时刺激表现出过度的炎症反应,主要是通过增加前列腺素E2(PGE2)的分泌,而PGE2由环氧合酶2(COX2)调节。因此,德国耶拿大学医院口腔正畸科、保守牙科和牙周病学系老年牙科课题组的一项研究旨在调查这种过度炎症是否由棕榈酸诱导的表观遗传改变介导。研究结果发表在 Cells 期刊题为“Palmitate-Triggered COX2/PGE2-Related Hyperinflammation in Dual-Stressed PdL Fibroblasts Is Mediated by Repressive H3K27 Trimethylation”。

为了阐明H3赖氨酸乙酰化(H3Kac)在调节同时受到机械压缩和细菌诱导的应激刺激的PA暴露HPdLFs的高炎症反应中的作用,实验首先检查了相关H3Kac调节因子(图1 a、b),其与PdL特性、牙周病和高脂血症有关。这些基因包括编码组蛋白乙酰转移酶(HATs)CREB结合蛋白(CBP),E1A结合蛋白p300(p300),赖氨酸乙酰转移酶8(KAT8)和核受体共激活因子3(NCOA3),以及编码组蛋白去乙酰化相关蛋白的基因,例如组蛋白去乙酰化酶1(HDAC1),HDAC2,HDAC3和对HDAC活性很重要的蛋白质,例如SIN3转录调节因子家族成员A(SIN3A)。
然而,定量表达分析显示,在PA处理下,无论是在编码HATs的基因(图1 a)还是HDACs 或 SIN3A的基因(图1 b),都没有生物学上相关的显著差异。因此,通过免疫荧光泛染色分析了H3K9/14/18/23/27ac(H3Kac)(图1 c、d),结果检测到,施加压缩力后H3K乙酰化水平增加,然而,由于PA暴露,没有观察到差异,这表明在这些条件下PA对H3Kac的影响相当小。
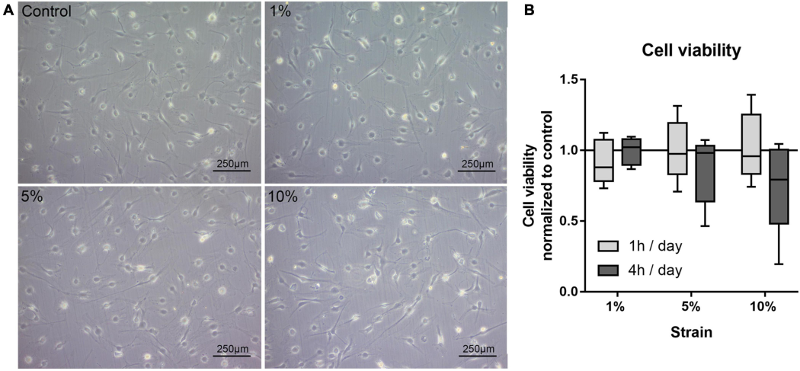
图1 棕榈酸对受牙龈假单胞菌LPS刺激的压缩下HPdLFs中的H3Kac没有影响。
考虑到PA也可以影响组蛋白甲基化的抑制形式,例如H3K27三甲基化(H3K27me3),接下来研究了这种修饰的改变是否有助于引发过度的炎症应激反应。实验对编码多梳抑制复合物2(PRC2)核心成分的基因进行了定量PCR,该基因对PdL功能很重要,包含EZH2及其高度相关的同源物EZH1、胚胎外胚层发育的抑制子(EED)、zeste 12的抑制子(SUZ12)。
数据显示,无论是压缩力还是脂肪酸暴露,均未检测到基因表达的相关变化(图2 a)。然而,PRC2的活性和特异性被证明受到核心成分的翻译后修饰的调节。因此,用定量免疫荧光分析了双刺激下HPdLFs中H3K27三甲基化水平(图2 b、c),发现施加额外的压缩力导致H3K27me3水平降低,然而,与对照组(82.68%±3.66)相比,PA培养组的变化(62.17%±4.24)明显较低。这说明,棕榈酸影响LPS 刺激的HPdLFs中压缩力诱导的H3K27me3降低。
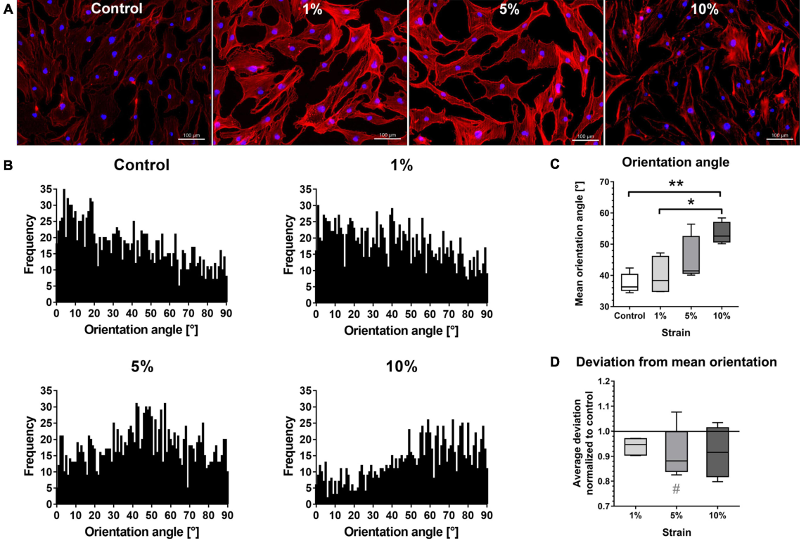
图2 在PA 暴露的HPdLFs中,K27三甲基化对双重刺激的反应较小。
随后研究了双应激PA培养中相应H3K27特异性组蛋白甲基转移酶(HMTs)活性的变化是否可能导致H3K27me3水平的改变,发现HMT活性在施加压缩力时降低,同样,在PA处理的HPdLFs中,这种趋势明显较弱。
为了评估H3K27三甲基化和HMT活性的这些微小变化是否与PA在双刺激HPdLFs中的促炎作用有关,实验用UNC1999抑制了PRC2、EZH1和EZH2的核心酶。浓度为1.00 μM 的UNC1999处理HPdLFs 后发现,H3K27相关组蛋白甲基转移酶(HMTs)的活性降低至23.23%±2.56%,尽管较低的抑制剂浓度也导致HMT活性的强烈抑制。
为了验证1.00 μM UNC1999对双应激下PA培养物中H3K27三甲基化的抑制作用,分析了HMT活性,在BSA对照和PA培养中,UNC1999处理导致HMT活性降低,证实了其在实验环境中的抑制作用。因此,与BSA对照相比,PA处理的HPdLFs对H3K27特异性HMT表现出相当的活性。
为了说明炎症过程,对单核细胞THP1细胞进行了粘附实验,与BSA对照相比,PA暴露导致双重刺激下HPdLFs的炎症反应增强。虽然UNC1999介导的EZH1和EZH2活性降低不会改变双刺激下BSA对照中贴壁THP1细胞的数量,但它将PA培养物中单核细胞的过度活化降低到与BSA对照相当的水平。
为了进一步研究UNC1999对炎症过程的影响,实验确定了COX2的表达,它似乎与PA诱导的双重刺激下HPdLFs的过度炎症反应有关。分析显示,PA培养物中的COX2表达显著高于BSA对照,这种上调的COX2转录被UNC1999抑制。为了确认COX2水平改变对相关细胞因子分泌的潜在影响,分析了双刺激下HPdLFs加UNC1999处理后上清中分泌的PGE2水平,结果支持先前在COX2表达背景下提出的假设,即双重刺激PA培养中PGE2的过量分泌被UNC1999所减弱。
总之,这些数据表明,双重刺激下PA培养物中H3K27me3的减少通过改变HMT活性可能导致HPdLFs的过度炎症反应,这可能是通过COX2/PGE2调节。
最后,实验研究了编码抗炎细胞因子白细胞介素10(IL-10,图3 a),已被证明可以调节COX2转录,其表达可能受到组蛋白修饰中脂肪酸依赖性变化的影响,发现在双刺激PA培养物中检测到IL10转录显著降低。
为了验证抑制H3K27三甲基化对IL10基因活性的影响,对与H3K27me3结合的DNA片段进行了染色质免疫沉淀。分析显示,在暴露于PA的双应激HPdLFs中,这种抑制组蛋白标记在两个IL10启动子相关侧(#1和#3)的水平增加(图3 b、c)。H3K27me3与IL10启动子位点(#1和#3)的增强关联被使用UNC1999而抑制的EZH1和EHZ2所抵消,结果与BSA对照相似(图3 b、c)。总之,与三甲基化H3K27的增强关联可能是PA处理双应激下HPdLFs中IL10表达降低的潜在原因,而IL10表达降低又可能导致这些条件下COX2/PGE信号传导增强。

图3 双应激下HPdLFs暴露于棕榈酸导致IL10启动子区域附近H3K27三甲基化降低,与IL10表达降低相关。
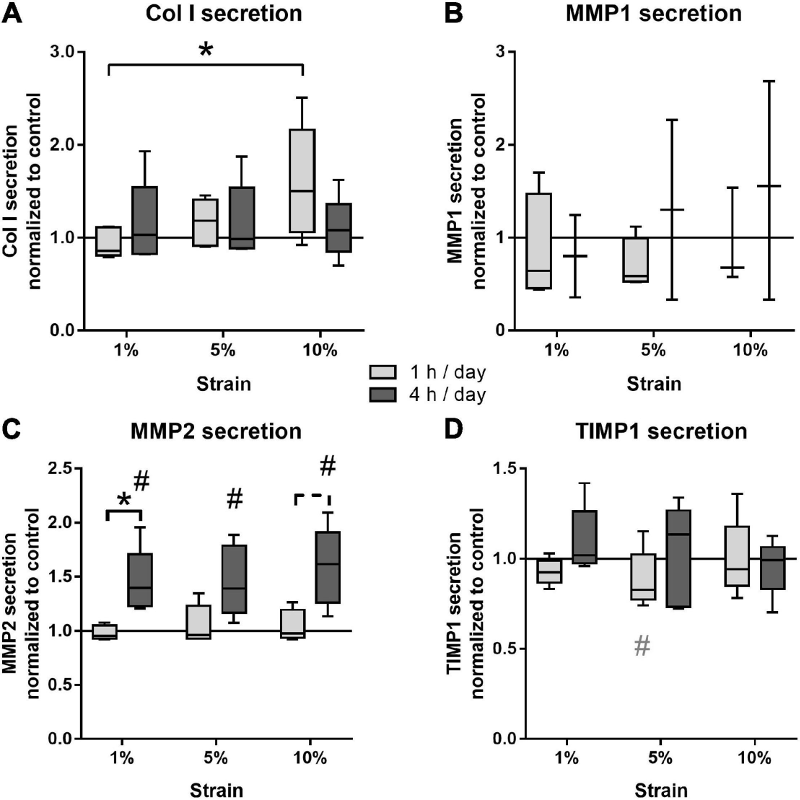
图4 图形概要
组蛋白的高度动态表观遗传修饰可以调节基因表达,以响应各种环境线索。该研究首次表明,肥胖相关的高脂血症状态可以通过影响其炎症反应的机械力影响细菌刺激的成纤维细胞中启动的表观遗传变化。因此,棕榈酸相关的对牙龈卟啉单胞菌LPS刺激的压缩下人PdLFs细胞的高炎症反应似乎是通过增强COX2抑制因子IL-10基因启动子附近的H3K27三甲基化而下调实现的。获得的结果强烈表明,代谢相关的变化在表观遗传调控中起着核心作用,这些调控是由致病性和机械应力引起的软硬组织重塑,为未来的靶向治疗提供了有希望的潜力。
参考文献:Schuldt L, Reimann M, von Brandenstein K, Steinmetz J, Döding A, Schulze-Späte U, Jacobs C, Symmank J. Palmitate-Triggered COX2/PGE2-Related Hyperinflammation in Dual-Stressed PdL Fibroblasts Is Mediated by Repressive H3K27 Trimethylation. Cells. 2022 Mar 10;11(6):955. doi: 10.3390/cells11060955. PMID: 35326406; PMCID: PMC8946768.
小编旨在分享、学习、交流生物科学等领域的研究进展。如有侵权或引文不当请联系小编修正。如有任何的想法以及建议,欢迎联系小编。感谢各位的浏览以及关注!
微信搜索公众号“Naturethink”,了解更多细胞体外仿生培养技术及应用。

新鲁汶大学的公报指出,如今抗生素耐药菌的出现给人类和医药带来了新 ...

根据“生物安全关键技术研发”重点专项评审工作安排,生物中心将于2 ...

为更好的向用户、潜在用户提供我们的产品,即日起推出如下活动:凡向 ...

2018年度国家科学技术奖提名工作已结束,国家科学技术奖励工作办 ...

据英国《自然·通讯》杂志日前发表的一篇医学论文报告,科学家发现了 ...

Naturethink祝愿伟大祖国繁荣昌盛,欣欣向荣! ...

“免疫系统在高血压中扮演了未曾预料的重要角色。”英国格拉斯哥大学 ...

“来一场中国制造的品质革命!”3月5日,李克强总理在政府工作报告 ...

我司自主研发产品,重视知识产权,已拥有多项专利证书! ...

美国侨报网近日刊文称,一项新出炉的研究警告称,即使是失眠一夜,也 ...
公司完成细胞张应变与压力综合培养仪器的研发; ...

Naturethink网站及微信内容逐步完善,敬请查阅! ...

为提高区域自主创新能力,推进区域科技创新体系建设,加大创新驱动区 ...

2018年春节将至,我司放假时间安排为:2月14日至2月21日, ...

澳大利亚和英国一项研究显示,对于几乎任何年龄段的人群而言,快走都 ...

2018年国家自然科学基金项目申请工作已开始,你准备好了吗? ...
壁面剪切应力(WSS),是单位面积上由血管表面流动的液体产生的接 ...

癌症在促进肿瘤表型表观遗传重编程和修饰的复杂组织微环境中发展。此 ...
